이 포스팅은 쿠팡 파트너스 활동의 일환으로, 이에 따른 일정액의 수수료를 제공받습니다.
비만과 제2형 당뇨병: 인슐린 저항성의 메커니즘
비만은 인슐린 저항성을 유발하는 핵심 요인이며, 이는 결국 제2형 당뇨병 발병의 주요 경로가 됩니다. 비만 상태에서는 지방조직에서 분비되는 비에스테르화 지방산, 글리세롤, 호르몬 및 염증성 사이토카인이 증가하고, 이들이 인슐린 저항성의 발달에 중요한 역할을 합니다. 이러한 상황에서 체지방 감소를 돕는 푸드올로지 콜레올로지컷 프로 다이어트 보조제와 같은 제품이 체중 관리에 도움을 줄 수 있습니다.
비만과 인슐린 저항성의 기본 원리
비만은 인슐린 저항성의 주요 유발 요인입니다. 인슐린 저항성은 인슐린의 혈당 흡수, 대사 또는 저장 효과에 대한 저항성을 의미합니다. 비만과 제2형 당뇨병에서 인슐린 저항성은 지방세포와 골격근에서의 인슐린 자극 포도당 수송 및 대사 감소와 간에서의 포도당 생성 억제 장애로 나타납니다.
인슐린 저항성은 인슐린 감수성(반응의 절반을 유도하는 인슐린 농도) 감소, 인슐린 반응성(인슐린 민감 조직에 대한 인슐린의 최대 효과) 감소, 또는 두 가지 모두의 형태로 나타날 수 있습니다.
비만에서 인슐린 저항성은 근본적으로 에너지 과잉 공급에 대한 피드백 조절 메커니즘을 나타냅니다. 세포 내 에너지 과잉으로 인한 미토콘드리아 과부하를 통제하기 위해 기질 흡수를 감소시키는 방향으로 작용합니다. 식단 조절과 함께 맛있닭 닭가슴살 스테이크 오리지널과 같이 단백질 함량이 높고 칼로리가 낮은 식품을 섭취하는 것이 에너지 밸런스 유지에 도움이 될 수 있습니다 [로켓프레시] 맛있닭 닭가슴살 스테이크 오리지널와 같은 제품을 활용할 수 있습니다.
인슐린 저항성의 분자적 메커니즘
JNK 신호 경로와 인슐린 저항성
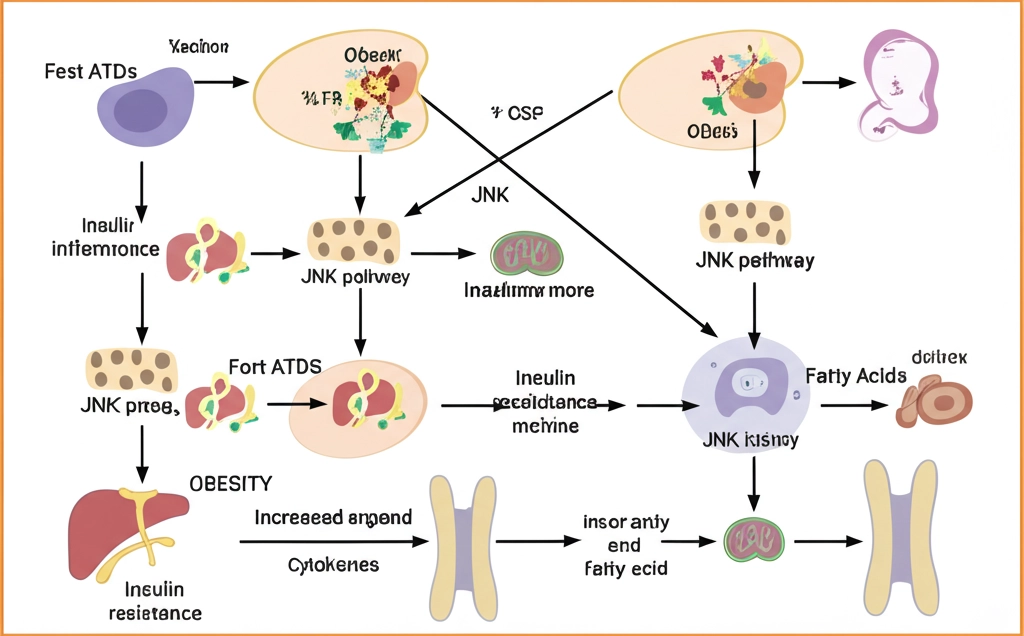
JNK(c-Jun N-terminal kinase)는 비만과 인슐린 저항성에서 가장 많이 연구된 신호 변환기 중 하나입니다. JNK는 MAPK(mitogen-activated protein kinase) 초가족의 일원이며, 성장 인자, 사이토카인, 스트레스 반응 및 기타 요인에 의해 활성화될 수 있습니다.
비만은 만성적인 낮은 수준의 염증을 유발하여 염증 인자와 유리 지방산(FFA)을 생성하고, 이는 인슐린 표적 세포에서 JNK 신호 경로를 활성화합니다. 예를 들어, TNFα와 같은 염증 인자는 수용체에 결합하여 JNK 신호 경로를 활성화하고, 이는 TNF 유전자 발현을 조절하여 악순환을 형성합니다.
비만 개인의 피하 지방 조직 생검에서 인산화된 JNK의 활성화 증가가 관찰되었으며, 이는 염증 반응을 더욱 증폭시키고 인슐린 신호 경로를 손상시킵니다. 간 특이적 JNK1 제거는 인슐린과 혈당 수준의 상당한 감소를 초래했으며, 간 Akt 인산화의 지속적인 증가를 보였습니다.
JNK는 IRS-1(인슐린 수용체 기질-1)의 세린 잔기를 인산화하여 인슐린 신호 전달을 억제합니다. 인산화된 IRS-1은 상위 인슐린 수용체와 상호작용하는 것을 방지할 뿐만 아니라 하위 PI3K를 활성화하지 못하게 됩니다.
세라마이드와 인슐린 신호 경로 방해
세라마이드는 인슐린 저항성에 중요한 역할을 합니다. 증가된 혈장 세라마이드 수준은 인슐린 저항성 및 대사 증후군 발병과 관련이 있습니다. 비만한 인슐린 저항성 동물 모델에서 증가된 팔미토일-CoA 가용성은 말초 조직에서 세라마이드의 de novo 합성과 스핑고미엘린 가수분해를 증가시킵니다.
세라마이드는 인슐린 자극 포도당 수송을 억제함으로써 인슐린 저항성에 중요한 역할을 합니다. 이는 주로 인슐린 신호 전달과 인슐린 매개 포도당 대사 자극에 필요한 세린/트레오닌 키나제인 Akt/PKB 경로를 억제함으로써 이루어집니다. 결과적으로 GLUT4 세포막 이동, 포도당 흡수, 그리고 글리코겐 합성이 모두 억제됩니다.
세라마이드는 두 가지 별개의 신호 분자인 단백질 포스파타제 2A(PP2A)와 단백질 키나제 C-제타(PKCζ)를 통해 Akt/PKB 신호 경로에 작용하여 포도당 대사를 방해합니다.
또한 세라마이드는 PKR/JNK 축을 활성화하여 IRS1의 인산화에 직접 관여하여 인슐린 수용체 신호 전달을 억제합니다.
PKC와 인슐린 수용체 조절
단백질 키나제 C(PKC)는 인슐린 저항성과 비만에서 중요한 역할을 합니다. 골격근의 인슐린 수용체 티로신 키나제 활성은 비만한 인슐린 저항성 환자에서 감소되어 있습니다.
알칼리성 포스파타제 처리는 비만 환자의 근육에서 in vitro 인슐린 자극 수용체 티로신 키나제 활성을 48% 증가시켰습니다. 이는 인슐린 수용체가 세린/트레오닌 잔기에서 과인산화되어 있음을 시사합니다.
인슐린 저항성 환자의 근육에서는 기저 상태에서 PKC-베타 단백질 함량이 증가하고, 인슐린 자극 조건에서는 막 관련 총 PKC 활성이 증가합니다. PKC-델타, -베타, -세타는 인슐린 처리에 반응하여 세포질에서 막 분획으로 이동합니다.
이러한 결과는 비만한 인슐린 저항성 환자의 골격근에서 인슐린 수용체 티로신 키나제 활성이 PKC에 의한 세린/트레오닌 인산화로 인해 감소되었을 가능성을 시사합니다.
IRS-1 Ser-1101 인산화와 S6K1 경로
S6K1(p70 ribosomal S6 kinase 1)은 IRS-1의 세린 잔기를 인산화하는 중요한 매개체입니다. IRS-1의 Ser-1101(사람 번호)은 S6K1에 의해 매개되는 인산화 부위로 확인되었습니다.
영양소 과부하와 비만 상태에서 IRS-1 Ser-1101의 인산화가 증가하고, 이는 PI3-키나제/Akt 신호 전달을 억제함으로써 인슐린 작용을 억제하는 데 관여합니다.
유전적으로 유도된 비만 마우스(db/db)의 간에서 S6K1 Thr-389와 IRS-1 Ser-1101 인산화가 현저히 증가했으며, 이는 급성 인슐린 자극에 의해 더욱 강화되었습니다. 반면, S6K1 결핍 마우스(S6K1-/-) 모델에서는 고지방식이로 유도된 인슐린 저항성과 IRS-1 Ser-1101 인산화 증가가 나타나지 않았습니다.
이러한 발견은 S6K1이 세포 자율적인 방식으로 작용하며, S6K1 제거(RNA 간섭에 의한)가 IRS-1 Ser-1101 인산화를 억제한다는 사실에 의해 더욱 뒷받침됩니다.
염증과 인슐린 저항성의 연관성
만성 염증과 인슐린 저항성
비만은 만성적인 낮은 수준의 염증을 유발하며, 이는 말초 조직의 인슐린 저항성을 유도하는 주요 메커니즘입니다. 마우스와 인간에서 지방조직의 염증은 세포 스트레스로 유도된 염증성 신호 경로의 활성화를 통해 인슐린 저항성을 유발합니다 산마을 간편 야채톡과 같은 제품이 효과적입니다. 염증 완화에 도움을 줄 수 있는 곰곰 열풍 서리태 볶음과 같은 건강 간식을 섭취하는 것도 좋은 방법입니다.
과도한 영양소, 지방분해, 당신생으로 인한 고지혈증과 고혈당은 미토콘드리아 기능 장애, 소포체 스트레스, 산화 스트레스를 유발하여 JNK와 IKKβ 같은 스트레스 반응성 신호 분자를 자극합니다.
JNK와 IKKβ 신호 경로는 IRS 세린-307 인산화 외에도 표적 조직에서 염증성 유전자 발현을 증가시켜 전신 염증을 증폭시킵니다. 또한, 포화 지방산과 장 유래 세균 지질다당류(LPS)는 TLR4(Toll-like receptor 4)에 결합하여 NF-κB와 JNK를 활성화하고, 염증과 인슐린 저항성을 매개합니다.
아디포카인과 인슐린 저항성
지방조직은 다양한 염증 촉진 및 억제 아디포카인을 분비하여 염증과 인슐린 저항성을 조절합니다. 비만한 인간과 설치류 모델에서 염증 촉진 아디포카인의 발현이 증가하여 인슐린 저항성을 유도합니다.
아디포카인의 불균형은 비만 유도 인슐린 저항성의 중요한 요인입니다. 렙틴과 아디포넥틴은 섭식 행동과 에너지 소비를 조절할 뿐만 아니라 염증 반응의 조절에도 관여합니다 닥터민스 식이섬유정1000과 같은 제품이 효과적입니다. 이러한 호르몬 균형을 맞추는 데 도움을 줄 수 있는 뉴트리나우 프롤린 모유 유산균과 같은 프로바이오틱스 제품을 섭취하는 것도 고려해볼 수 있습니다.
비만 연구에서는 염증 촉진 및 억제 아디포카인 발현의 불균형이 비만 유도 인슐린 저항성의 주요 원인 중 하나임을 제시합니다.
SOCS3와 골격근 인슐린 저항성
SOCS3(Suppressor of cytokine signaling-3)는 시토카인 신호 전달 억제제로, 비만한 인간의 골격근에서 발현이 증가합니다. SOCS3는 시상하부에서 렙틴 신호 전달과 지방조직 및 간에서 인슐린 신호 전달을 억제합니다.
골격근은 에너지 소비와 전신 인슐린 감수성을 조절하는 중요한 조직이지만, 이 조직에서 SOCS3의 생리적 중요성은 충분히 연구되지 않았습니다. 이를 조사하기 위해 골격근에서 특이적으로 SOCS3이 제거된 마우스(SOCS3 MKO)가 개발되었습니다.
SOCS3 MKO 마우스는 고지방식이로 유도된 비만 정도가 정상 마우스와 유사했지만, 인슐린 증가와 인슐린 저항성 발달로부터 보호되었습니다. 이는 개선된 골격근 인슐린 수용체 기질 1(IRS1)과 Akt 인산화로 인한 것으로, 이는 증가된 골격근 포도당 흡수로 이어졌습니다.
골격근에서 SOCS3를 억제하는 치료법은 비만 관련 포도당 불내성과 인슐린 저항성을 개선하는 데 효과적일 수 있습니다. 체지방 감소를 돕고 혈당 관리에 도움을 줄 수 있는 그린몬스터 다이어트 스페셜 2 가르시니아 900과 같은 제품을 섭취하는 것도 하나의 방법이 될 수 있습니다 랩앤뷰티 뉴티 차전자피 식이섬유 6g 30포도 좋은 선택입니다.
세포 기관 기능 이상과 인슐린 저항성
미토콘드리아 기능 이상과 인슐린 저항성
비만 상태에서 미토콘드리아 과활성화가 인슐린 저항성을 유발할 수 있다는 여러 증거가 있습니다:
1. β-산화에서 미토콘드리아 기능 향상이 인슐린 저항성을 초래했으며, 약리학적 억제제(에토목시르 또는 옥스페니신)로 β-산화를 억제하면 설치류에서 지질 유도 인슐린 저항성이 예방되었습니다.
2. 감소된 미토콘드리아 기능은 더 나은 인슐린 감수성과 관련되고, 증가된 미토콘드리아 기능은 트랜스제닉 마우스에서 인슐린 저항성을 유도했습니다.
3. 건강한 인간 대상에서, 지질 주입으로 유도된 인슐린 저항성은 미토콘드리아 β-산화 증가와 관련되었습니다.
인슐린 감작제(TZDs, 메트포민, 베르베린 등)는 모두 세포 모델에서 ATP 생산의 미토콘드리아 기능을 억제했습니다. 이와 더불어, 꾸준한 체중 변화를 확인하기 위해 앳플리 T3 디지털 LED 체중계를 사용하는 것도 체중 관리 여정을 효과적으로 추적하는 데 도움이 될 수 있습니다 상쾌한 식이섬유 젤리 사과맛 / 난소화성말토덱스트린와 같은 제품을 활용할 수 있습니다.
이러한 증거들은 비만에서 미토콘드리아 과활성화가 인슐린 저항성을 유발할 수 있다는 주장을 뒷받침합니다.
비만에서 인슐린에 민감한 세포의 ATP 생산은 에너지 수요와 관계없이 증가하는데, 이를 "미토콘드리아 과열"이라고 부릅니다. 과열은 미토콘드리아에 대한 기질 과잉 공급으로 인해 발생하여 추가적인 ATP 생산을 초래합니다. ATP 과잉 생산은 AMPK 억제, mTOR 유도, 고인슐린혈증, 글루카곤 증가, 미토콘드리아 기능 장애와 같은 여러 메커니즘을 통해 전신 인슐린 저항성에 기여합니다.
소포체 스트레스와 인슐린 저항성
소포체 스트레스는 주로 산화 스트레스 증가를 통해 인슐린 저항성의 병인에 중요한 역할을 합니다. 이는 특히 비알코올성 지방간 질환에서 볼 수 있습니다.
비만으로 인한 만성 저등급 염증은 TNF-α를 활성화시켜 ERK 및 JNK를 통해 PPARγ의 전사 활성을 억제합니다. PPARγ 리간드는 유도성 산화질소 합성효소, 매트릭스 메탈로프로테이나제, 인터루킨과 같은 다양한 염증 관련 유전자의 발현을 억제합니다.
이러한 행동은 비만 관련 인슐린 저항성과 관련될 수 있는데, 지방 조직의 대식세포 축적과 유전자 발현이 비만 유도 인슐린 저항성의 병인에 중요한 역할을 하는 것으로 입증되었기 때문입니다.
인슐린 저항성에서 제2형 당뇨병으로의 진행
고인슐린혈증과 베타세포 기능
비만에서 고인슐린혈증은 인슐린의 과잉 생산 또는 감소된 제거에서 비롯됩니다. 혈장 인슐린 수준은 인슐린 생산률과 인슐린 제거률의 균형에 의해 결정됩니다.
비만에서 체중 증가 동안 췌장 섬의 베타세포 기능이 향상되고 세포 수가 증가합니다. 이러한 변화는 지방산/포도당에 의한 베타세포 자극과 관련이 있습니다.
베타세포에서의 렙틴 저항성은 베타세포에 의한 인슐린 과잉 생산에 기여할 수 있습니다. 렙틴은 베타세포에서 인슐린 생산을 억제합니다. 베타세포 특이적 렙틴 수용체 녹아웃 마우스에서 이러한 억제가 사라지고 고인슐린혈증이 관찰되었습니다. 이 마우스는 먼저 고인슐린혈증을 보인 다음 인슐린 저항성을 발전시켰습니다.
이 연구는 렙틴 저항성 마우스 모델에서 고인슐린혈증에 대한 탁월한 설명을 제공합니다.
인슐린 저항성과 당 대사 장애
지방독성은 인슐린 저항성과 췌장 베타세포 기능 장애에 역할을 합니다. 순환 지질 수준의 증가와 지방산 활용 및 세포 내 신호 전달의 대사적 변화는 근육과 간의 인슐린 저항성과 관련되어 있습니다.
새로운 PKC 경로와 JNK-1 경로와 같은 다양한 경로가 지방독성이 간과 근육과 같은 지방이 아닌 조직 기관에서 인슐린 저항성을 유발하는 메커니즘으로 관여합니다.
미토콘드리아 기능 장애는 인슐린 저항성의 병인에 역할을 합니다. 소포체 스트레스는 주로 산화 스트레스 증가를 통해 인슐린 저항성의 병인에 중요한 역할을 하며, 특히 비알코올성 지방간 질환에서 볼 수 있습니다.
내장 지방과 인슐린 저항성은 모두 심장대사 위험을 증가시키며, 지방독성은 이러한 연관성의 병태생리학에 중요한 역할을 하는 것으로 보입니다.
결론
비만과 제2형 당뇨병의 연관성에서 인슐린 저항성은 핵심적인 병태생리학적 메커니즘입니다. 본 보고서에서는 JNK 신호 경로, 세라마이드와 지방독성, PKC 활성화, S6K1 매개 IRS-1 인산화, 염증 과정, SOCS3의 역할, 미토콘드리아 및 소포체 스트레스 등 다양한 분자적 메커니즘을 살펴보았습니다.
이러한 메커니즘들은 복잡하게 상호작용하며 인슐린 신호 전달 경로를 방해하고, 인슐린 저항성을 유발하며, 궁극적으로 제2형 당뇨병으로 이어집니다. 또한 지방조직에서 분비되는 아디포카인과 염증성 사이토카인이 인슐린 저항성의 발달과 유지에 중요한 역할을 합니다.
이러한 메커니즘에 대한 이해는 비만 관련 인슐린 저항성 및 제2형 당뇨병을 효과적으로 치료하기 위한 새로운 치료법 개발에 중요한 기반을 제공합니다. 특히, JNK 억제제, 세라마이드 합성 억제제, 골격근 SOCS3 억제, 미토콘드리아 기능 조절 등은 유망한 치료 타겟이 될 수 있습니다.
비만과 인슐린 저항성의 복잡한 관계를 이해하는 것은 대사 질환의 예방 및 치료를 위한 포괄적인 접근 방식을 개발하는 데 필수적입니다. 향후 연구는 이러한 메커니즘들 간의 상호작용을 더 깊이 탐구하고, 특정 대사 상태 및 생활 방식 중재에 대한 더 정확한 바이오마커를 정의하며, 자연적 또는 합성 약물을 통한 타겟 치료법을 개발하는 데 중점을 두어야 할 것입니다.
댓글
댓글 쓰기